
Prof. Dr. Wilfried Brade (Hannover/Dummerstorf) und Dr. Edwin Brade (Paretz)
Hochleistende Holstein-Kühe – zunehmend leider auch immer häufiger als "Turbokühe" bezeichnet – seien zu stark auf Leistung getrimmt, so lautet ein häufiger Vorwurf der Öffentlichkeit. Sogar von Qualzucht ist teilweise die Rede (1, 6, 42).
Die Zuchtbranche wischt diese Bedenken gerne mit dem Hinweis beiseite, die meisten Verbraucher hätten ohnehin nur geringe Kenntnisse von der modernen Milcherzeugung. Aber diese (einseitige) Reaktion löst keine Probleme; weder das Imageproblem, das mit der Debatte auf die Branche zurollt, noch die (in einigen Betrieben tatsächlich bestehenden) Probleme im Hinblick auf die Tiergesundheit.
Sicher gibt es viele Betriebe, die mit Herdenleistungen von 11.000 kg und mehr erfolgreich sind, deren Kühe größtenteils gesund sind und eine hohe Lebensleistung erreichen. Aber es gibt eben auch die anderen, die mit dem Ausfüttern und Managen solcher Leistungen nicht (mehr) zurechtkommen.
Stoffwechsel- und Eutererkrankungen, Lahmheiten und schlechte Fruchtbarkeit gehören eben auch zur Realität und resultieren in einer insgesamt zu geringen Nutzungsdauer der Tiere. Am Ende bleibt als Antwort auf die häufig gestellte Frage: "Ob die deutsche oder internationale Holsteinzucht ein Problem hat?", oft nur die (unzureichende) Antwort: Es kommt darauf an:
" ... das Herdenmanagement ist wohl entscheidend ...".
Wie lässt sich also auf die (sich anbahnende) öffentliche Diskussion reagieren?
Eine gesetzliche Beschränkung der Leistungszucht ist sicher nicht die gewünschte Lösung. Allerdings forderte selbst der deutsche Tierärztetag im Oktober 2015 in seiner Erklärung die Politik auf, Zuchtziele zu überprüfen und gegebenenfalls zu korrigieren, um Erkrankungen und Schäden zu minimieren. Es ist also höchste Zeit für die Zuchtbranche, zu handeln (1, 6, 9, 11, 42).
Die Holstein-Zuchtverbände sind gefragt, die Anteile der Merkmale in der (Gesamt-) Zuchtwertschätzung zu überprüfen und gegebenenfalls neue Merkmale in den Zuchtprozess zu integrieren (3, 8, 11, 12, 13, 19, 23, 24, 29, 34, 44, 51).
Hier rückt nun auch die Futteraufnahme – speziell im ersten Laktationsdrittel hochleistender Holstein-Kühe – in den züchterischen Blickpunkt (9).
Die konsequente Erhöhung der Milchleistung basierte vorzugsweise auf einer Selektion nach einer hohen Einsatzleistung. Ungeachtet der begrenzten Futteraufnahmekapazität – vor allem zu Beginn der Laktation – hat dies zu einer dramatischen Zunahme der negativen Energiebilanz (NEB) geführt. Eine lang andauernde und starke NEB wird als ein bedeutender Risikofaktor für verschiedene Erkrankungen oder Fruchtbarkeitsstörungen angesehen (5, 9, 15, 26, 29, 31, 39). Jüngste Zuchtstrategien betrachten daher nicht nur die Einsatzleistung, sondern auch den Zeitpunkt des Laktationspeaks oder die Dauer des Leistungsplateaus (9, 40, 46, 47, 51).
Die Futteraufnahme von Milchkühen ist ein wichtiges Merkmal, das in aktuellen deutschen Zuchtprogrammen (noch) nicht berücksichtigt wird. Dies liegt in erster Linie daran, dass in der Praxis die Futteraufnahme tierindividuell nur mit großem Aufwand zu bestimmen wäre. Nur in wenigen Landesversuchsanstalten und Forschungsinstituten lässt sich die Futteraufnahme tierindividuell und täglich bestimmen.
Ob jedoch neuere genetisch-züchterische Ansätze in Form der genomisch gestützten Selektion zukünftig eine Alternative darstellen, ist derzeit in Diskussion.
Bei der Kuh besitzt das Laktationsstadium eine überragende Bedeutung für die Futteraufnahme (2, 3, 27, 28, 32, 65).
Im ersten Drittel der Laktation ist das Futteraufnahmevermögen im Vergleich zu späteren Laktationsabschnitten niedriger (3, 32, 33, 52, 59, 65). Aber gerade in dieser Phase ist der Bedarf an Energie und Nährstoffen für den Erhalt und die Milchproduktion am höchsten (Abbildung 1).
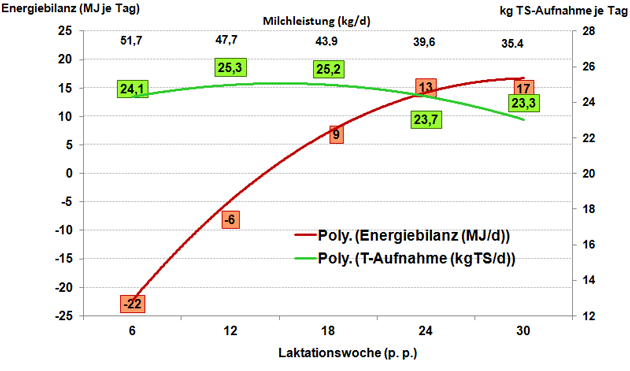
Abbildung 1: Energiebilanz (MJ/d) und Futteraufnahme (kg TS/d) bei hochleistenden Kühen bis zur 30. Laktationswoche.
Quelle: Eigene Grafik.
Entsprechend kann die hochleistende frühlaktierende Milchkuh ihren Nährstoff- und Energiebedarf nicht in ausreichendem Maß über das aufgenommene Futter decken. Ein Teil der nötigen Energie kann aber über die Mobilisierung von Körperreserven bereitgestellt werden.
Die Mobilisierung von Körpersubstanz (Glycogen, Fett und Protein) im Zeitraum der Geburt ist ein evolutionär manifestierter Prozess, der bei allen Säugetieren, wenngleich in unterschiedlichem Ausmaß, auftritt. Nur wenn weit vor der Geburt größere Mengen an Körperreserven angelegt wurden, können diese im geburtsnahen Zeitraum auch mobilisiert werden. Kühe mit einem hohen Körperkonditionsindex sind entsprechend in der Lage, mehr Fett im geburtsnahen Zeitraum zu mobilisieren. Der Prozess der Mobilisierung beginnt bereits mehrere Wochen vor dem Kalben und ist bei der Milchkuh circa eine Woche vor der Kalbung durch einen Anstieg der Fettsäurekonzentration im Blutplasma sowie einem Rückgang der Futteraufnahme nachweisbar. Die mobilisierten Energiereserven vor der Kalbung dienen vor allem zur Gewinnung von Energie, der Versorgung des Fötus, dem Aufbau der Milchdrüse und der Produktion von Kolostrum.
Es ist zu beobachten, dass das Ausmaß angelegter Körperfettreserven die Futteraufnahme bereits vor der Kalbung negativ beeinflusst. Kühe, die bereits vor der Kalbung ein hohes Maß an Körperfettreserven angelegt haben, besitzen im Vergleich zu ihren schlankeren Stallgefährtinnen eine niedrigere Futteraufnahme. Dieser Unterschied bleibt oft während des gesamten ersten Laktationsdrittels, in der die Futteraufnahme kontinuierlich ansteigt, manifest.
Kühe mit einer hohen Körperfettmobilisierung weisen in der Frühlaktation auch einen erhöhten Milchfettgehalt oder Milchfett/Milcheiweiß-Quotienten auf. Die Hinzuziehung von Körperreserven zur Milchfettsynthese stellt einen energiesparenden Mechanismus für die Kuh dar. So würde im Vergleich zur Milchfettsynthese aus körpereigenen Fettsäuren für die de novo Synthese von Milchfett aus Acetat, welches im Pansen aus dem aufgenommenen Futter gebildet wird, ein zig-faches an Energie aufgewendet werden müssen. Andererseits hat das Ausmaß der Mobilisierung einen entscheidenden Einfluss auf die Stoffwechselbelastung der Tiere und die daraus resultierende Stoffwechselgesundheit und das Wohlbefinden (25, 29, 39, 40).
Ferner können weitere energiebenötigende Anpassungsvorgänge (wie die Adaptation des Pansens und des Intestinaltrakts an energiereichere Rationen mit weniger Struktur, der verstärkte Abfluss von Blut-Calcium in die Milchdrüse und eine generelle Schwächung des Immunsystems) die Stoffwechselgesundheit beeinträchtigen (5, 26, 29, 31).
Eine übermäßige Körperfettmobilisation spiegelt sich in stark erhöhten Konzentrationen an nicht-veresterten Fettsäuren (non-esterified fatty acids, NEFA) im Blut wieder, welche ihrerseits zu einer erhöhten Ketonkörperproduktion und Fettakkumulation in der Leber führen können. Eine schwerwiegende Fettanreicherung, die sogenannte Fettleber, kann störend auf Stoffwechselfunktionen der Leber wirken (5, 15, 16, 45, 48, 51).
Bei zu fetten Tieren ist infolge einer erhöhten Mobilisation von Körpersubstanz und der gleichzeitig relativ geringeren Futteraufnahme auch mit einer verstärkten Neigung zu Stoffwechselstörungen zu rechnen.
Die NEB ist neben dem Rückgang der Körpermasse (KM) auch durch eine Abnahme der Körperkondition (Boniturnoten, BCS) charakterisiert (16, 45, 52, 60). Die Reduktion des Körpergewichts geht dabei über den Gewichtsverlust, der durch das Fruchtwasser und das Gewicht des Kalbes sowie die Rückbildung des Uterus erklärt werden kann, weit hinaus.
Bereits frühere Untersuchungen zeigten, dass in den ersten acht Wochen nach dem Kalben im Mittel über 45 kg Körpermasse (bei einer hohen tierindividuellen Schwankungsbreite von etwa 20 bis 90 kg) mobilisiert werden können (55). Die Tendenz, vorhandene Köperreserven in immer kürzerer Zeit zu mobilisieren, hat sich mit zunehmendem Selektionsdruck auf höhere Einsatzleistung in den vergangenen Jahren weiter verstärkt (22).
Da das Futteraufnahmevermögen der Kuh in der Frühlaktation begrenzt ist, erfordert die rasch steigende Milchleistung in diesem Zeitraum eine hohe Energiedichte in der Gesamtration, die oftmals durch hohe Kraftfutteranteile realisiert wird. Ein hoher Kraftfutteranteil kompromittiert allerdings Leistung, Gesundheit und ernährungsphysiologisch-relevante Aspekte wie:
Zusätzliche Beachtung erfordert speziell auch – wie bereits genannt – die Proteinversorgung der Hochleistungskuh in der Phase der intensiven Mobilisation von Körpermasse (KM).
Ein Kilogramm KM besitzt einen Energiegehalt von etwa 21 MJ NEL; sein Eiweißgehalt entspricht aber "nur" einem Äquivalent von 320 g Rohprotein (46, 47). Mit anderen Worten: Aus einem Kilogramm mobilisierter KM kann die Kuh etwa 7,2 kg Milch (aus energetischer Sicht), aber gleichzeitig nur 3,8 kg Milch aus Blickrichtung der Proteinbereitstellung bilden.
Die benötigten Proteine werden bevorzugt aus der Skelettmuskulatur mobilisiert. Eine intensive KM-Mobilisation geht mit einem Proteindefizit einher, das nun zusätzlich fütterungsseitig zu beachten bleibt (47).
Beispiel: Eine Kuh baut täglich etwa 1,6 kg KM in der fünften Laktationswoche ab. Dieser KM-Abbau liefert Energie für rund 11,5 kg Milch pro Tag; aber reicht "nur" für rund 6 kg Milch pro Tag hinsichtlich der Proteinbereitstellung. Etwa 460 g unabbaubares Futterprotein (UDP, engl.: undegradable protein) müssen der Kuh zusätzlich über die Ernährung bereitgestellt werden (das heißt 84 g UDP je kg "Milchdifferenz"), sollen die oben genannten 11,5 kg Milch pro Tag vollständig realisiert werden (46, 47).
Sind die Körperreserven allerdings erschöpft und weist die Kuh gleichzeitig weiterhin ein deutliches Energiedefizit (zum Beispiel aufgrund einer unzureichenden Trockenmasseaufnahme) auf, sinkt die Milchleistung ebenso wie der Milchfett- und -eiweißgehalt drastisch ab.
Die Futteraufnahme (FA) hängt wie die Leistungseigenschaften von genetischen und nicht-genetischen Faktoren ab (Abbildung 2).

Abbildung 2: Vereinfachtes Schema zur Regulation der Futteraufnahme beim Rind.
Quelle: Eigene Darstellung; in Anlehnung an (43).
Erstkalbinnen nehmen prinzipiell weniger Trockensubstanz auf als ältere Kühe (27, 28, 43, 57). Ihre Aufnahmekapazität liegt im Alter von zwei Jahren bei etwa 80 Prozent gegenüber ihren älteren Stallgefährtinnen. Aufgrund der allgemein geringeren Milchleistung erstlaktierender Kühe geraten diese entsprechend nicht so stark in eine NEB.
Generell ist zu beachten, dass die Futteraufnahme der Tiere physikalischen, chemischen und hormonellen Steuerungsmechanismen unterliegt.
(28) haben umfassende Gleichungen zur Vorhersage der FA – unter Berücksichtigung des Laktationsstadiums – abgeleitet. Diese Gleichungen sind komplex und nur über entsprechende, computergestützte Rationsprogramme vorteilhaft anzuwenden.
Neben dem Tier ist die Qualität des Futters von entscheidender Bedeutung für die Höhe der FA (Abbildung 2). (28) nutzen deshalb auch den Energiegehalt des Grobfutters bei der Vorhersage der FA. Daneben bestimmen Geruch und Geschmack des Futters die Höhe der FA.
Zu vermerken ist, dass die Regulation der FA entsprechend der Milchleistung im ersten Drittel der Laktation noch wenig ausgeprägt ist. Während zu Anfang der Laktation die FA (erfasst über die Trockenmasseaufnahme) im Mittel um nur etwa 0,1 kg Trockensubstanz (TS) je Kilogramm Milch steigt, erhöht sich die Futteraufnahme zu Ende der Laktation im Mittel um 0,2 kg TS je Kilogramm Milch (28). Dies kann dadurch erklärt werden, dass für die Synthese der Milch im Zeitraum der Frühlaktation auch körpereigene Reserven herangezogen werden.
Entsprechend verringert sich die Lebendmasse in der Frühlaktation, die im Vergleich zur Entwicklung der Futteraufnahme gegenläufig ist (3, 16, 28, 37, 45, 52). Erst in der zweiten Laktationshälfte frisst die Kuh entsprechend ihrer Milchleistung (Energieabgabe).
Somit sind im Betriebsmanagement alle Maßnahmen zu ergreifen, die eine hohe FA zu Beginn der Laktation begünstigen.
Folgende Punkte sind bereits während der Trockenstehzeit und beim Laktationsstart zu beachten:
Bei der Regulation der Futteraufnahme sind sowohl kurzfristige als auch langfristige Signalnetzwerke beteiligt, die darauf abzielen, dem Tier Hunger- und Sättigungsgefühle zu vermitteln. Kurzfristig wirken vor allem postingestive Signale zur Beendigung einer Mahlzeit. Daran beteiligt sind vor allem Mechano- und Chemorezeptoren, die die Füllung des Pansens oder die Konzentration der unterschiedlichen Nährstoffe detektieren (49).
Die Weiterleitung der Signale erfolgt dann über das vegetative Nervensystem zum Hirnstamm, wo sie umgeschaltet und zum Hypothalamus projiziert werden. Der Hypothalamus ist der zentrale Integrator für Signale, die die Futteraufnahme bestimmen. Hier werden zum Beispiel auch weitere Reize aus anderen Hirnregionen wie optische Eindrücke, Gerüche und Geschmackswahrnehmungen integriert. Darüber hinaus kennt man Hormone, die in Abhängigkeit der Nahrungsaufnahme aus dem Magen-Darm-Trakt in die Blutbahn abgegeben werden und so zum Hirnstamm oder direkt zum Hypothalamus gelangen (Abbildungen 3 und 4).
Während zahlreiche gastrointestinale Polypeptide ein Sättigungsgefühl erzeugen, wurde dem Ghrelin-Hormon, das beim Wiederkäuer in der Pansenwand und im Dünndarm gebildet wird, eine futteraufnahmesteigernde Wirkung zugesprochen (66). Jüngste Untersuchungen belegen jedoch, dass Ghrelin für die Futteraufnahmesteigerung in der Frühlaktation eher eine untergeordnete Bedeutung besitzt und dessen Wirkung auf den Energiestoffwechsel als viel bedeutsamer einzuschätzen ist (7).
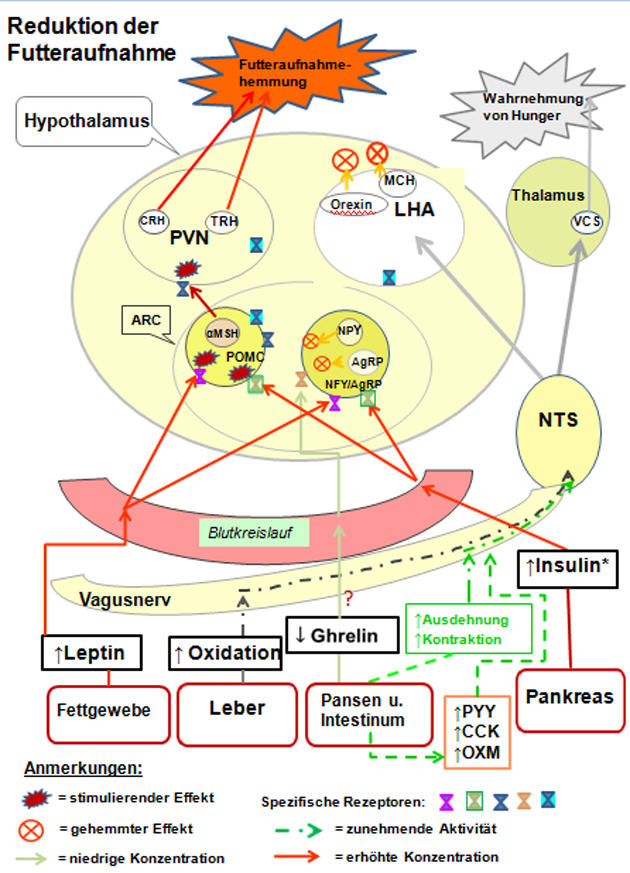
Abbildung 3: Vereinfachte schematische Darstellung beteiligter Signalnetze sowie Aktivitäten im Hypothalamus bei der Futteraufnahme.
Quelle: Eigene, modifizierte Darstellung in Anlehnung an (50).
Anmerkungen: * Glucose hat beim Wiederkäuer keinen Sättigungseffekt.
AgRP = Agouti-related protein; ein endogener Melanocortinantagonist; experimentell verursacht es Fresssucht und Fettsucht, Hyperglykämie und Hyperinsulinismus (bei Mäusen);
ARC = Nucleus arcuatus; ein Kern des Hypothalamus;
CCK = Cholecystokinin; ein Peptidhormon des Magen-Darm-Trakts;
CRH = Corticotropin-releasing Hormone, ein Polypeptid, das im Hypothalamus (im PVN) gebildet wird;
LHA = Lateraler Hypothalamus (engl. Lateral Hypothalamic Area); er regt bei einer Stimulation den Hunger an (nach der dualen Hypothalamustheorie des Hungers);
MCH = Melanin-konzentrierendes Hormon; gilt als einer der wichtigsten körpereigenen "Appetitanreger"; es soll auch das Wachstum und die Zahl der insulinherstellenden Zellen in der Bauchspeicheldrüse fördern (Mensch);
NPY = Neuropeptid Y, es ist neben der zentralnervösen Steuerung des Hungers wohl auch an der Insulinfreisetzung beteiligt;
NTS = Nucleus tractus solitarii; ist für die Geschmackswahrnehmung (Geschmackskern) zuständig, sowie bei der Verarbeitung von viszeralen Afferenzen von Chemo- oder Barorezeptoren (Dehnungsrezeptoren); gleichzeitige Beteiligung am Atem- oder Würgereflex;
OXM = Oxyntomodulin; ein Peptidhormon, das im Dünndarm (dort von den sogenannten L-Zellen während der Nahrungsaufnahme) freigesetzt wird und eine Rolle bei der Nahrungsaufnahmeregulation spielt;
PVN = Nucleus paraventricularis; ein Kerngebiet im Hypothalamus;
PYY = Peptide tyrosine tyrosine; ein Peptid, das (beim Menschen) den Appetit zu reduzieren vermag;
TRH = Thyreotropin Releasing Hormon (Thyreoliberin oder auch Protirelin genannt); ein Peptidhormon, das im Hypothalamus gebildet wird;
VCS = Visceral sensory complex; er ist beteiligt an der Verwaltung des zur Hirnrinde gerichteten Informationsflusses (Thalamus).
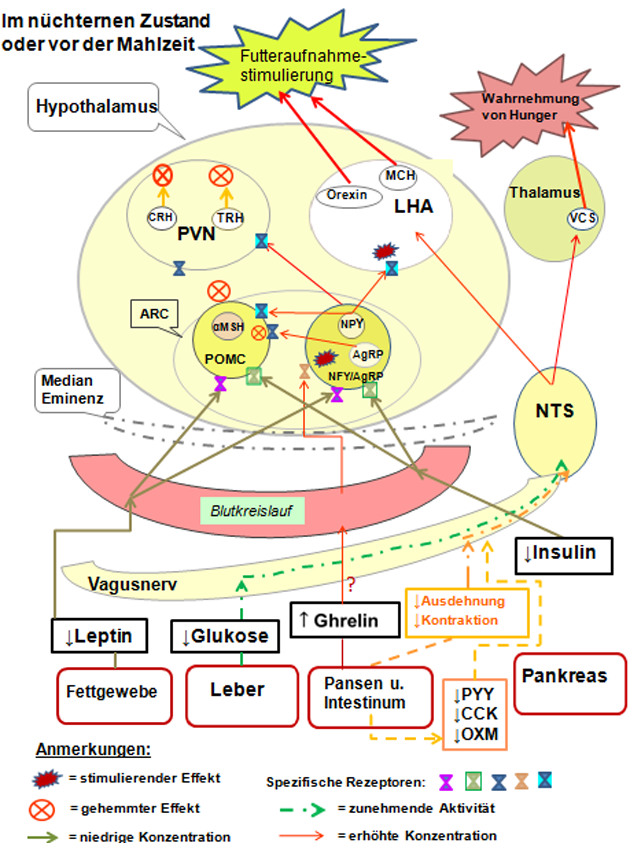
Abbildung 4: Vereinfachte schematische Darstellung beteiligter Signalnetze sowie Aktivitäten im Hypothalamus im nüchternen Zustand.
Quelle: Eigene, modifizierte Darstellung in Anlehnung an (50).
Anmerkungen:
AgRP = Agouti-related protein; ein endogener Melanocortinantagonist; experimentell verursacht es Fresssucht und Fettsucht, Hyperglykämie und Hyperinsulinismus (bei Mäusen);
ARC = Nucleus arcuatus; ein Kern des Hypothalamus;
CCK = Cholecystokinin; ein Peptidhormon des Magen-Darm-Trakts;
CRH = Corticotropin-releasing Hormone, ein Polypeptid, das im Hypothalamus (im PVN) gebildet wird;
LHA = Lateraler Hypothalamus (engl. Lateral Hypothalamic Area); er regt bei einer Stimulation den Hunger an (nach der dualen Hypothalamustheorie des Hungers);
MCH = Melanin-konzentrierendes Hormon; gilt als einer der wichtigsten körpereigenen "Appetitanreger"; es soll auch das Wachstum und die Zahl der insulinherstellenden Zellen in der Bauchspeicheldrüse fördern (Mensch);
NPY = Neuropeptid Y, es ist neben der zentralnervösen Steuerung des Hungers wohl auch an der Insulinfreisetzung beteiligt;
NTS = Nucleus tractus solitarii; ist für die Geschmackswahrnehmung (Geschmackskern) zuständig, sowie bei der Verarbeitung von viszeralen Afferenzen von Chemo- oder Barorezeptoren (Dehnungsrezeptoren); gleichzeitige Beteiligung am Atem- oder Würgereflex;
OXM = Oxyntomodulin; ein Peptidhormon, das im Dünndarm (dort von den sogenannten L-Zellen während der Nahrungsaufnahme) freigesetzt wird und eine Rolle bei der Nahrungsaufnahmeregulation spielt;
PVN = Nucleus paraventricularis; ein Kerngebiet im Hypothalamus;
PYY = Peptide tyrosine tyrosine; ein Peptid, das (beim Menschen) den Appetit zu reduzieren vermag;
TRH = Thyreotropin Releasing Hormon (Thyreoliberin oder auch Protirelin genannt); ein Peptidhormon, das im Hypothalamus gebildet wird;
VCS = Visceral sensory complex; er ist beteiligt an der Verwaltung des zur Hirnrinde gerichteten Informationsflusses (Thalamus).
Die mit der Nahrung aufgenommenen Nährstoffe werden nach Verdauung und Absorption dem Intermediärstoffwechsel der einzelnen Organe zugeführt, wo sie unter Entstehung von ATP (Adenosintriphosphat) oxidiert werden. Da der Großteil des Futters eines Wiederkäuers aus Kohlenhydraten besteht, gewinnt der Wiederkäuer den Großteil seiner Energie aus Kohlenhydratoxidation (KOX). Entsprechend verringert sich die KOX, wenn keine Nahrung aufgenommen wird. Jüngere Untersuchungen belegen den Zusammenhang zwischen der Aufnahme einzelner Mahlzeiten und der KOX bei spätlaktierenden Milchkühen (21).
Die langfristigen Signalnetzwerke sind darauf ausgerichtet, die Energiebilanz auszugleichen und damit das Körpergewicht konstant zu halten. Als wichtigstes Hormon, das Auskunft über die vorhandenen Fettreserven gibt, gilt das Leptin. Dieses Adipokin wir vom Fettgewebe gebildet und spiegelt direkt proportional die Körperfettmassen wider.
Insulin wird nicht zuletzt aufgrund seiner steten Wirkung auf die Glucosehomeostase ebenfalls eine Langzeitwirkung auf die Futteraufnahme zugeschrieben. Beide Hormone besitzen prinzipiell eine futteraufnahmereduzierende Wirkung. Der Rückgang der Leptin- und Insulinkonzentration von der Spätträchtigkeit bis zur Frühlaktation kann somit als futteraufnahmesteigernde Signalgebung angesehen werden.
Da Kühe mit der höheren Körperkondition auch höhere Plasmaleptinkonzentrationen aufweisen, ist es verständlich, dass sie im Vergleich zu Tieren mit geringerer Körperkondition die niedrigere Futteraufnahme aufweisen.
Die FA ist ein Merkmal, das in aktuellen Milchrinderzuchtprogrammen (noch) fehlt. Trotz der großen Bedeutung der Futteraufnahme zur Realisierung gewünschter Leistungen und des bekannt hohen Beitrages der Futterkosten an den Produktionskosten ist die FA bisher nicht in Zuchtprogramme integriert worden.
Der Verzicht auf die FA in aktuellen Zuchtzielen bei Milchrindern ist vor allem auf das Fehlen von tierindividuellen Routine-Daten zur Futteraufnahme in Praxisbetrieben zurückzuführen.
Verfügbare Daten zur FA bei Milchkühen umfassen vorrangig Versuchsanstellungen oder Daten aus speziellen Versuchsherden.
Die zwischenzeitlich zahlreich vorliegenden (weltweiten) Versuchsergebnisse bestätigen, dass nicht nur für die Milchleistung, sondern auch für die Futter- oder Energieaufnahme eine deutliche genetische Variation zwischen den Milchkühen vorhanden ist. Allerdings ist die Heritabilität (h2, Erblichkeit) der FA in der Regel geringer als die der Milchleistung, das heißt hier sind auch die möglichen Selektionserfolge prinzipiell geringer einzuschätzen (Tabelle 1). Auch sind die Heritabilitäten im Laktationsverlauf nicht konstant (4, 14, 37, 56, 65). Gleichzeitig bestätigen weiterführende quantitativ-züchterische Auswertungen, dass die genetischen Beziehungen (rg) zwischen der Milchleistung und der FA generell nur im mittleren Bereich liegen (Tabelle 1).
| Tabelle 1: Studien zur Heritabilität (h2) der Futteraufnahme und genetische Beziehungen (rg) zwischen Futteraufnahme und Milchleistung | ||||||
|---|---|---|---|---|---|---|
| Quelle | Tiermaterial | Tieranzahl (n) | h2 | genetische Korrelation (rg) zwischen FA und Milch (Merkmal) | Fütterungs-system | Bemerkungen (FA: zugehörige Definition des Merkmals) |
| HOOVEN et al., 1972 (gekürzt) | Jungkühe (HF) | 425 | 0,241 0,262 | 0,521 (FCM) 0,862 (FCM) | Raufutter betont | FA: Energieaufnahme 131 - 60 Tage p. p. 21 - 300 Tage p. p. (Gesamtlaktation) |
| GRAVERT, 1985 | Jungkühe (vorrangig HF) | 96 (Lak- tationen); Zwillings-studie | 0,16 | 0,12 (FCM) (1. bis 20. Woche p. p.) | TMR mit ¾ Maissilage, ¼ Kraftfutter, 1 kg Heu | FA: Futterenergie-aufnahme (MJ NEL) |
| VAN ARENDONK et al., 1991 | Jungkühe (vorrangig HF) | 360 | 0,461 0,312 | 0,611 (FPCM) 0,612 (FPCM) (105 Tage p. p.) | Raufutter ad libitum, 6 kg Kraftfutter | FA: 1Raufutter-aufnahme (kg TS/d), 2Energieaufnahme (MJ ME/d) |
| LEE et al., 1992 | Jungkühe (HF) | 1.266 | 0,27 | 0,95 (Milch menge, kg); unterschiedliche Zeiträume | Kraftfutter: leistungs-bezogen; Raufutter: ad libitum | FA: Energieaufnahme Milchmenge (kg): 0 - 24 Wochen p. p.; Energieaufnahme: 8 - 16 Wochen p. p. |
| JENSEN et al., 1995 | Jungkühe (verschiedene Rassen) | 295 | 0,16 | 0,50 (ECM) (250 Tage p. p.) | TMR, ad libitum | FA: tägliche Energieaufnahme |
| VEERKAMP und BROTHERSTONE, 1997 | Jungkühe (HF) | 410 | 0,43 | 0,34 (Milch-Kg) 0,49 (Eiw.-Kg) | TMR, ad libitum | FA: TS-Aufnahme (kg/d) in den ersten 26 Wochen p. p. |
| VEERKAMP et al., 1995 | Jungkühe + Kühe | 377 | 0,36 | 0,44 (Milchenergie) | TMR, ad libitum | FA: ME-Aufnahme (MJ/d); in den ersten 26 Wochen p. p. |
| HÜTTMANN et al., 2009 (gekürzt) | Jungkühe (HF) | 289 | 0,061 0,112 0,113 0,114 | 0,441 (ECM) 0,422 (ECM) 0,663 (ECM) 0,904 (ECM) | TMR, ad libitum | FA: TS-Aufnahme (kg/d); 1 11. - 30. Tag p. p., 2 31. - 60. Tag p. p., 3 61. - 90. Tag p. p., 4 91. - 120. Tag p. p. |
| BUTTCHEREIT et al., 2012 (gekürzt) | Jungkühe (HF) | 682 | 0,05 bis 0,20* | -0,01 bis 0,57 (in verschied- Laktations-abschnitten)* | TMR, ad libitum | FA: TS-Aufnahme (kg/d); * 15. - 150. Laktationstag p. p. |
| LIINAMO et al., 2012 (gekürzt) | Jungkühe (Nordische Rote) | 291 | 0,23 (2. - 30. Woche) | -0,101 (ECM) 0,272 (ECM) 0,693 (ECM) 0,754 (ECM) | TMR
| FA: TS-Aufnahme (kg/d); 1 2. Woche p. p., 2 5. Woche p. p., 3 10. Woche p. p., 4 15. Woche p. p. |
| SPURLOCK et al., 2012 (gekürzt) | Jungkühe + Kühe (HF) | 402 | 0,34* | 0,61 (ECM) | TMR, ad libitum | FA: TS-Aufnahme (kg/d); * nur 1. Monat nach dem Kalben hier dargestellt |
Anmerkungen: TS = Trockenmasse, d = Tag, FA = Futteraufnahme, ECM = energiekorrigierte Milch, FCM = fettkorrigierte Milch; FPCM = korrigierte Milch nach Fett- und Proteingehalt.
Die FA sollte auf eine Selektion nach höherer Milchleistung reagieren; die zugehörigen genetischen Effekte dürften jedoch nur begrenzt sein (Tabelle 1).
Diese begrenzten Selektionseffekte lassen nachteilige Auswirkungen auf die NEB (Intensität, Dauer) erwarten, falls weiter einseitig auf höhere Milchleistung selektiert wird.
Bestätigt wird der beschriebene Zusammenhang auch in der Literatur. So hat (22) eine Veränderung der Milchleistung der Änderung der Futteraufnahme bei US-amerikanischen Milchkühen gegenübergestellt: Im Zeitraum von 1980 bis 2003 erhöhte sich die Milchleistung um über 50 Prozent, die Futteraufnahme jedoch lediglich um circa 25 Prozent, das heißt die Milchleistung und damit der Energiebedarf der Milchkühe veränderten sich rasanter als die FA.
Da sich die Merkmalszusammenhänge zwischen FA, Milchleistung und Energiebilanz (EB) im Laktationsverlauf deutlich verändern, sind die genetischen Beziehungen zwischen der FA zu Beginn der Laktation und in späteren Laktationsabschnitten nicht identisch (rg ≤ 1,0 – Tabelle 2).
| Tabelle 2: Geschätzte Heritabilitäten (in der Diagonalen) für die Futteraufnahme (TS-Aufnahme) und genetische Korrelationen zwischen den Merkmalswerten, erfasst zu unterschiedlichen Zeitpunkten in der ersten Laktation (Versuchsbetrieb) | |||||
|---|---|---|---|---|---|
| DIM* | |||||
| DIM | 11 | 30 | 80 | 130 | 180 |
| 11 | 0,37 (0,04) | 0,96 (0,01) | 0,62 (0,06) | 0,39 (0,07) | 0,32 (0,08) |
| 30 | 0,34 (0,03) | 0,81 (0,03) | 0,60 (0,05) | 0,41 (0,07) | |
| 80 | 0,28 (0,03) | 0,92 (0,01) | 0,61 (0,05) | ||
| 130 | 0,27 (0,03) | 0,84 (0,03) | |||
| 180 | 0,37 (0,04) | ||||
Quelle: (56).
Anmerkung: * DIM = Tage in Milch p. p..
Für den Genetiker sind diese veränderten Merkmalszusammenhänge ein deutlicher Beleg dafür, dass in verschiedenen Laktationsabschnitten verschiedene Gene unterschiedlich wirksam sind.
Auf den ersten Blick scheint die Einbeziehung der Futteraufnahme in ein Zuchtprogramm eine relativ einfache Sache zu sein. Die Tücke liegt jedoch im Detail: die notwendige differenzierte und korrekte Bewertung der FA innerhalb verschiedener Laktationsabschnitte; insbesondere diejenige im ersten Laktationsdrittel.
Die klassische Selektionstheorie erlaubt die Quantifizierung möglicher merkmalsspezifischer Zuchtfortschritte bei differenzierter Auslese der Zuchttiere (9, 12, 20, 35, 59).
Nachfolgend werden die zu erwartenden Selektionserfolge in den ersten 100 Laktationstagen von Holstein-Jungkühen – bei unterschiedlicher Auswahl der eingesetzten Bullen1 (Kuhväter) – aufgezeigt
Die verwendeten genetisch-züchterischen Populationsparameter zur Ableitung notwendiger Selektionsindizes sowie zu erwartende Erfolge sind in den Tabellen 3 und 4 zusammengestellt.
| Tabelle 3: Populationsgenetische Eingangsparameter zur Bestimmung des Selektionserfolges bei differenzierter Zuchtwahl; Basis: Jungkühe in den ersten 100 Tagen p. p. (praktischer Betrieb) | ||||||
|---|---|---|---|---|---|---|
| Merkmal | Mittelwert (x) | phäno- typische Streuung (Sp) | Variations- Koeffizient (v in Prozent) | Herita- bilität (h²) | genetische Korrelationen (rg) | |
| FA | KM | |||||
| ECM | 3.200 | 480 | 15 | 0,25 | 0,32 | 0,26 |
| FA | 18,8 | 1,504 | 8 | 0,15 | identisch | 0,40 |
| mittlere KM | 600 | 42 | 7 | 0,40 | 0,40 | identisch |
Anmerkungen: ECM = energiekorrigierte Milch (kg/Kuh), FA = Futter-Trockenmasseaufnahme (kg TS/d), KM = Körpermasse (kg) im Mittel der ersten 100 Tage p. p..
| Tabelle 4: Erwartete Selektionserfolge in der Nachkommengeneration (ΔG) bei differenzierten Selektionsrichtungen der Vatertiere | ||||
|---|---|---|---|---|
| Selektionsrichtung: (ökonomische Gewichtsver-hältnisse (ECM:FA:KM) im Zuchtziel) | Selektionserfolge in der ersten Nachkommengeneration (ΔG) | Relative Höhe der Futter-energieaufnahme gegenüber dem Bedarf in den ersten 100 Tagen (p. p.) in Prozent* | ||
| ECM | FA** | KM*** | ||
| ausschließlich auf ECM (1:0:0) | + 411,9 | + 0,271 | + 12,3 | 88,8 |
| kombiniert mit Selektion auf Milch und FA (1:100:0) | + 404,8 | + 0,417 | + 16,0 | 89,5 |
| kombiniert mit hoher Bewertung der FA und leicht negativer Bewertung der Körpermasse (1:500:-3) | + 330,5 | + 0,673 | + 10,5 | 92,3 |
| kombiniert mit sehr hoher Bewertung der FA und negativer Bewertung der Körpermasse (1:500:-5) | + 321,1 | + 0,623 | + 2,3 | 92,5 |
Anmerkungen: * Bedingung: Futterenergiekonzentration im Mittel 7,2 MJ NEL/kg täglich verzehrte Futtertrockenmasse; ** FA = tägliche Futter-Trockenmasseaufnahme (kg TS/d); *** mittlere Körpermasse in den ersten 100 Tagen p. p..
Die genetisch-züchterischen Veränderungen in den Nachkommenschaften sind bei differenzierter Auslese der Besamungsbullen deutlich verschieden.
Zunächst wurde – zwecks Abschätzung (maximal) möglicher Selektionserfolge für die Milchleistung (ECM) – eine einseitig spezialisierte Auslese der Kuhväter ausschließlich auf Milchleistung vorausgesetzt (erste Zeile in Tabelle 4); anschließend eine gleichzeitige Auslese der Vatertiere bei (sehr) differenzierter Gewichtung der Milchleistung (ECM) und FA abgebildet (zweite Zeile in Tabelle 4). Schließlich wurde zusätzlich auch die mittlere Körpermasse der Kühe berücksichtigt.
Bei einseitig konsequenter Selektion der Kuhväter ausschließlich auf Milchleistung kann die Milchleistung der Töchter um circa 412 kg Milch in der Nachkommengeneration verbessert werden; Gleichzeitig fällt die Energiebilanz (EB) – trotz eines gleichzeitig positiven Zuchtfortschrittes auch in der FA (+ 0,27 kg TS-Aufnahme pro Tag) – auf 88,8 Prozent.
Da zusätzlich auch eine enge positive Beziehung zwischen FA und mittlerer Körpermasse (KM) für die Jungkühe im ersten Laktationsdrittel, aufgrund der Hinweise von (28, 57, 59), vorausgesetzt wurde (Tabelle 3), erhöht sich bei betonter Selektion auf Futteraufnahme gleichzeitig die Körpermasse der Kühe (Tabelle 4).
Da mit Zunahme der KM bekanntermaßen wiederum der Erhaltungsbedarf steigt, wurde schließlich – bei sehr intensiver Selektion auf FA – zusätzlich eine negative Bewertung der Körpermasse vorausgesetzt (dritte und vierte Zeile in Tabelle 4).
Die Gesamtergebnisse bestätigen, dass eine einseitige Selektion auf hohe Milchleistung im ersten Laktationsdrittel die EB der Milchkuh verschlechtert. Gleichzeitig verdeutlichen die dargestellten Zusammenhänge, dass eine weitere unerwünschte Entwicklung in der Energiebilanz (Ausmaß, Dauer) hochleistender Milchkühe im ersten Laktationsdrittel künftig eine positive Bewertung der Futteraufnahme im Selektionsprozess erfordert; unter gleichzeitiger Berücksichtigung weiterer Merkmale (hier: Körpermasse und -kondition).
Selektionsempfehlungen für die Holstein-Züchtung in dieser Phase der Laktation sind somit nicht trivial.
Bei konsequent zusätzlicher und positiver Einbeziehung des Merkmals "Futteraufnahme" im ersten Laktationsdrittel in den künftigen Selektionsprozess – fortlaufend angewendet über weitere nachfolgende Generationen – sollte somit die weitere, schnelle Ausdehnung der NEB, im Vergleich zur einseitigen Selektion auf Milchleistung, klar begrenzt werden können!
Die Etablierung der genomisch gestützten Selektion eröffnete die Möglichkeit, neue Merkmale in Zuchtprogrammen zu nutzen (3, 4, 8, 10, 61). Die Futteraufnahme ist ein solches Merkmal; bei notwendiger differenzierter Bewertung speziell dieses Merkmals im Laktationsverlauf.
Allerdings sind hier große Bullennachzuchten mit zugehörigen Merkmalsinformationen – wie sie routinemäßig für die Milchleistung zur Verfügung stehen – als Referenzgruppe auch zukünftig nicht verfügbar.
Deshalb wird aktuell sehr intensiv – über den internationalen Austausch von zugehörigen Versuchs- und Praxisdaten – an der gemeinsamen Erstellung eines großen Trainingssets zur genomischen Selektion gearbeitet.
Im Projekt "Global Dry Matter-Initiative" arbeiten zurzeit 15 Partner aus zehn Ländern zusammen. Ihr Ziel ist: Bildung einer gemeinsamen Referenzpopulation mit Holstein-Tieren, die genotypisiert sind und zusätzlich Informationen zur Trockensubstanzaufnahme (TS-Aufnahme) aufweisen (3, 4, 61, 64).
(4) zeigen, dass es möglich ist, internationale Daten zu kombinieren, um Zuchtwerte zu generieren. Korrekterweise müssen aber Genotyp-Umwelt-Interaktionen, vor allem für Weideproduktionssysteme, berücksichtigt werden (Abbildung 5).
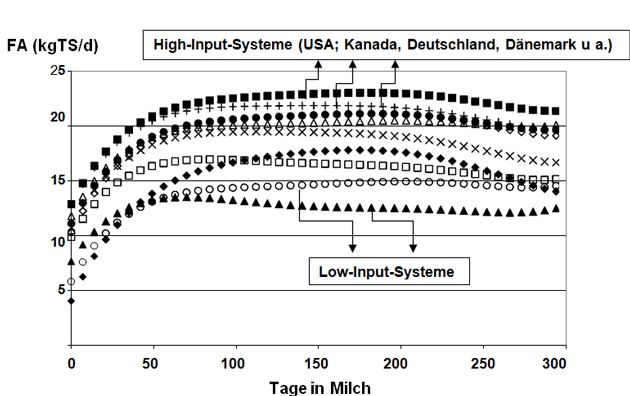
Abbildung 5: Futteraufnahme (in kg TS/d) von Jungkühen in der ersten Laktation in verschiedenen Studien unter differenzierten Umweltbedingungen.
Quelle: Eigene Grafik, nach Angaben von (4).
Die Höhe und der Verlauf der FA lässt deutliche Einflüsse des Produktionssystems erkennen (Abbildung 5). Es sollte deshalb auch nicht überraschen, dass die genetischen Korrelationen zwischen der Futteraufnahme, die in verschiedenen Regionen/Produktionssystemen ermittelt wurden, deutlich verschiedenen sind und klar von eins (rg ≤ 1) abweichen (Tabelle 5).
| Tabelle 5: Genetische Korrelationen (rg) zwischen der FA in verschiedenen Produktionssystemen; internationaler Datensatz | |
|---|---|
| Region | EU: High-Input |
| EU: Low-Input | 0,84 (0,14) |
| Nordamerika | 0,76 (0,21) |
| Weidensysteme (Low-Input) | 0,33 (0,20) |
Quelle: (4).
Unabhängig von der genannten internationalen Zusammenarbeit, die dringend benötigte weitere Resultate zur FA erwarten lässt, dürfte die Verfügbarkeit tierindividueller Daten zur FA in praxi immer ein limitierender Faktor für genaue Zuchtwertschätzungen von Zuchttieren (Besamungsbullen) sein.
Es gibt deshalb weltweit ein zusätzliches Interesse an indirekten Vorhersagekennwerten, um langfristig die Genauigkeit der Zuchtwerte für die FA zu verbessern.
So gibt es den (zusätzlichen) Ansatz, ausgewählte Exterieurmerkmale mit der Milchleistung und/oder der Körpermasse zur Vorhersage der FA zu kombinieren (16, 17, 18, 44, 59, 60, 62, 63, 64, 65).
Andere mögliche Ansätze basieren auf:
Regelmäßige Erfassungen von tierindividuellen Aktivitätsdaten zum Fressverhalten sind gleichzeitig sehr informativ und hilfreich, um spezifische Krankheiten oder auch die Brunst vorherzusagen. Auch liefern veränderte Fressmuster zusätzliche Hinweise auf veränderte soziale Hierarchien (zum Beispiel nach Neugruppierung von Milchkühen).
Eine ungenügende Futteraufnahme, insbesondere zu Beginn der Laktation, ist am Futterzustand (Körperkondition) der Kühe leicht visuell erkennbar.
Eine einseitige Leistungserhöhung zu Laktationsbeginn lässt eine weitere Zunahme der NEB erwarten. Die NEB in der Frühlaktation begrenzt bei hochleistenden Kühen somit bereits aktuell die weitere Leistungssteigerung im ersten Laktationsdrittel (Abbildung 6).

Abbildung 6: Zahl der Tage mit Energieunterversorgung in Abhängigkeit von der Leistungshöhe; Rationstyp: Grassilage bestimmt.
Quelle: Eigene Berechnungen.
Da neben der Milchleistung die FA nach dem Kalben das Ausmaß der NEB mitbestimmt, sind alle Fütterungs- und Managementmaßnahmen – beginnend bereits am Ende der Vorlaktation oder in der Trockenstehperiode – darauf auszurichten, dass die Kühe eine hohe FA p. p. erreichen.
Um die Entstehung von Krankheiten wie beispielsweise Ketose zu verhindern, kommt es auf zwei entscheidende Faktoren an (5, 25, 26, 39, 46):
Die Futteraufnahme hochleistender Kühe bedarf, wie gezeigt werden konnte, aus genetisch-züchterischer als auch ernährungsphysiologischer Sicht im ersten Laktationsdrittel einer hoch positiven Bewertung. Eine ausschließliche Betrachtung der Futtereffizienz im Sinne einer ökonomischen Verbesserung ist nicht angezeigt, da dabei das Tierwohl und etwaige Kosten für die Behandlung frühpostpartaler Erkrankungen nicht berücksichtigt werden.
Die Vermeidung einer Überkonditionierung während der vorhergehenden Laktation oder in der Trockenstehperiode, ein geringer Umweltstress und ein hoher Kuhkomfort sind hier zusätzlich zu nennen.
Die Erfassung der Futteraufnahme wird eine Herausforderung bleiben, zumal die Merkmalserfassung zu unterschiedlichen Laktationszeitpunkten erforderlich ist.
Aus der Blickrichtung der Züchtung bietet sich jedoch nun die genomische Selektion an. Aktuell kann die Genauigkeit genomischer Vorhersagen für die FA bereits mit circa r2 ≈ 0,4 angegeben werden (44).
Die Tatsache, dass es bald möglich sein wird, genomische Zuchtwerte für die FA (mittels kombinierter Referenzpopulationen) abzuschätzen (44, 64), begründet die Dringlichkeit, wie am besten diese neuen Informationen im Zuchtziel (demnächst) zu verwenden sind, ohne dass es zu unerwünschten und nachteiligen Reaktionen bei den Nachkommen kommt (Abbildung 7).

Abbildung 7: Nutzung einer internationalen Referenzpopulation zur möglichen Vorhersage von genomischen Zuchtwerten für die Futteraufnahme in verschiedenen Laktationsabschnitten.
Quelle: Eigene Darstellung.
Das Projekt Global Dry Matter-Initiative basiert bereits auf über 10.000 Holstein-Tieren, die genotypisiert sind und zusätzlich weitere Informationen zum Beispiel zur Trockensubstanzaufnahme (TS-Aufnahme) innerhalb verschiedener Laktationsabschnitte aufweisen (3, 4); zwecks Ableitung von Schätzformeln für genomisch begründete Zuchtwerte für die Futteraufnahme (Abbildung 7).
(3, 4) zeigen auch, dass es möglich ist, internationale Daten erfolgreich zu kombinieren, um Zuchtwerte zu generieren (Abbildung 5).
Die Tatsache, dass es in Kürze möglich sein wird, hinreichend genaue genomische Zuchtwerte (mittels international kombinierter Referenzpopulationen) abzuschätzen, erfordert eine klare Bewertung, wie am besten die Futteraufnahme im Zuchtziel zukünftig zu verwenden ist, ohne dass es zu unerwünschten Reaktionen bezüglich der Tiergesundheit und des Tierwohls kommt.
Der vorliegende Beitrag soll zur Beantwortung dieser Frage im Kreise der praktizierenden Tierärzte dienen; bewußt abweichend von rein wirtschaftlichen Überlegungen in der Milcherzeugung.
Die künftige Züchtung bei hochleistenden Milchrindern erfordert die zusätzliche Etablierung und Nutzung des Selektionskriterium "Futteraufnahme" zur Vermeidung einer weiteren Ausdehnung der negativen Energiebilanz im ersten Laktationsdrittel aber auch zur Verbesserung der Futtereffizienz im zweiten und dritten Laktationsdrittel.
Die Leistungen der Milchkühe sind in Deutschland – speziell in den zurückliegenden Jahren – aufgrund einer erfolgreichen Selektion auf höhere Milchleistung bei gleichzeitig verbesserter Fütterung, Haltung und tierärztlicher Betreuung rasant gestiegen. Mittlere Herdenleistungen von über 11.000 kg Milch/Kuh/Jahr sind heute keine Seltenheit mehr.
Eine korrekte Bewertung der Futteraufnahme (FA) erfordert eine differenzierte Bewertung dieses Merkmals im Laktationsverlauf, da einerseits die FA in verschiedenen Laktationsabschnitten unterschiedlich genetisch determiniert ist und andererseits die Beziehungen zur Milchleistung variieren.
Die FA reagiert indirekt auf eine einseitige Selektion bezüglich einer höheren Milchleistung. Modellkalkulationen auf der Basis der modernen Selektionstheorie zeigen jedoch, dass eine einseitige Selektion auf höhere Milchleistung, speziell im ersten Laktationsdrittel, die Energiebilanz (EB) der Milchkuh weiter verschlechtert.
Das Vermeiden einer weiteren unerwünschten Entwicklung der NEB (negative Energiebilanz) sowohl in Hinblick auf das Ausmaß als auch die Dauer im ersten Laktationsdrittel erfordert künftig eine positive Bewertung der Futteraufnahme im Selektionsprozess; unter gleichzeitiger Berücksichtigung weiterer Merkmale innerhalb dieses Laktationsabschnittes (zum Beispiel Körpermasse). Im zweiten und dritten Drittel der Laktation, in denen die Tiere sich allgemein in einer ausgeglichenen Energiebilanz befinden, ist die genetisch-züchterische Bewertung der FA deutlich verschieden vom ersten Laktationsdrittel vorzunehmen.
Da zukünftig der Tiergesundheit und dem Tierwohl sowie auch der Bioethik bei der Erzeugung von Lebensmitteln tierischer Herkunft – im Hinblick auf die Akzeptanz der modernen Tierhaltung in der Öffentlichkeit – eine viel stärkere Bedeutung zukommt, sollte die Einbeziehung der Futteraufnahme in den Zuchtprozess bei Milchkühen zwecks Vermeidung einer weiteren Zunahme der NEB, speziell im Hochleistungsbereich, intensiver gefordert werden.
In Germany – especially in recent years –, dairy cow performance has increased rapidly due to successful selection towards higher milk yield and improved feeding, husbandry and veterinary care. Average herd performances of over 11,000 kg of milk/cow/year are no longer uncommon.
The correct evaluation of feed intake (Futteraufnahme, FA) requires a differentiated assessment of this feature during lactation given that, on the one hand, feed intake is genetically determined differently in different lactation periods and, on the other, genetic relations to milk yield vary.
Feed intake reacts indirectly to an unilateral selection towards higher milk yield. However, model calculations based on the modern theory of artificial selection show that unilateral selection towards higher milk production, especially in the first trimester of lactation, further deteriorates the dairy cow’s energy balance (EB). To avoid further undesired effects on the negative energy balance (NEB) – where both extent and duration during the first trimester of lactation are concerened – feed intake during the selection process must be evaluated positively while considering other features (such as body mass, for instance) within this phase of lactation. In the second and third phases of lactation, during which the animals’ energy status is normally balanced, feed intake, in terms of both genetic and breeding aspects, needs to be evaluated in ways significantly different from those in the first lactation phase.
Given the fact that – where public acceptance of modern animal husbandry is concerned – animal health and welfare as well as bioethics in the production of foods of animal origin will gain in importance in the future, the inclusion of feed intake in the breeding process of dairy cows (in order to avoid a further increase in NEB) should be requested much more vigorously.
Le rendement des grandes laitières en Allemagne – surtout dans les dernières années – à augmenté de manière très significative étant donnée la sélection unilatérale vers plus de performance laitière qui va de pair avec des meilleures conditions d’alimentation, d’élevage et de soins vétérinaires. Des performances de troupeaux d’une moyenne au dessus de 11.000 kg de lait/vache/an ne sont plus incommuns.
Une évaluation correcte de l’alimentation (Futteraufnahme, FA) demande une évaluation différenciée de ce facteur au cour de la période de lactation étant donné que, d’un côté, l’alimentation est génétiquement déterminée de manières différentes au cours des périodes de lactations variées et de l’autre les rapports avec la performance laitière varient.
L’alimentation réagit indirectement à une sélection unilatérale visant à une performance laitière augmentée. Pourtant, des calculations modèles basées sur la théorie de sélection moderne montrent qu’une sélection unilatérale visant à plus de performance laitière, surtout au premier trimestre de lactation, détériore le bilan énergétique négatif (BEN) de la vache laitière. Pour éviter que le BEN continue à développer de manière indésirable, et en ce qui est de leur étendu et au niveau de leur durée au cours du premier trimestre de lactation, une évaluation positive de l’alimentation durant le processus de sélection sera demandée tout en considérant également d’autres facteurs (comme la masse corporelle, par exemple) durant cette même période de lactation. Pendant le deuxième et le troisième trimestre de lactation, pendant lesquels le bilan énergétique des animaux est généralement équilibré, les aspects de l’alimenation au niveau génétique et au niveau de l’élevage doivent être évalués de manière très différente de l’évaluation au premier trimestre de lactation.
Etant donné que dans l’avenir, la santé et le bien-être des animaux ainsi que la bioéthique dans le cadre de la production d’aliments d’origine animale jouerant un plus grand rôle en vue de l’acceptance publique de l’élevage moderne, l’inclusion de l’alimentation dans le processus d’élevage des vaches laitières pour éviter une croissance du BEN, surtout au niveau des animaux de haute performance, devra être réclamée plus vivement.
1 Vorausgesetzt wird hier eine konventionelle Auslese der Kuhväter auf der Basis einer klassischen Indexselektion; aufbauend auf eine systematische Nachkommenprüfung und Verfügbarkeit von je 15 Töchterleistungen zur merkmalsspezifischen Zuchtwertschätzung (mittels der genomischen Selektion lassen sich erfahrungsgemäß circa 15 Töchterleistungen abbilden (8, 10, 20, 64). Die Selektionsintensität (i) bei den Vätern beträgt i = 2,06, das heißt nur fünf Prozent der geprüften Besamungsbullen werden als Kuhväter weiter genutzt.
Prof. Dr. Wilfried Brade
Tierärztliche Hochschule Hannover (TiHo)
zurzeit: Leibniz-Institut für Nutztierbiologie Dummerstorf (FBN)
Wilhelm-Stahl-Allee 2
18196 Dummerstorf
Email: brade@fbn-dummerstorf.de
Dr. Edwin Brade
Rinderspezialberater
14669 Ketzin/Paretz